
Interazioni tra disturbo antropico ed organizzazione spaziale nel capriolo (Capreolus capreolus Linnaeus, 1758) in ambiente mediterraneo. Home range e caccia al cinghiale.
- Pietro Cobre
- 19 Febbraio 2014
- Ambiente & Natura, Zoologia
- 0 Comments
Interazioni tra disturbo antropico ed organizzazione spaziale
nel capriolo (Capreolus capreolus Linnaeus, 1758)
in ambiente mediterraneo.
Home range e caccia al cinghiale.
Dott. Pietro Cobre (Zoologo) e Dott. Paolo Montanaro (Zoologo)
1. INTRODUZIONE
Il presente studio si inserisce a conclusione del progetto di reintroduzione del capriolo italico sui Monti della Tolfa realizzato dalla Provincia di Roma nel 2001 e nel 2002 e ha lo scopo di analizzare le interazioni tra il disturbo antropico nell’area di rilascio e l’organizzazione spaziale nel nucleo di individui reintrodotti in ambiente mediterraneo.
Il disturbo antropico di maggior risalto che si è riscontrato nell’area di studio è rappresentato dalla presenza, nel periodo che va dal 1 novembre al 31 gennaio, della caccia praticata nei confronti del cinghiale. Questo tipo di caccia viene eseguita con la tecnica tradizionale della braccata.

Dalla letteratura esaminata si è potuto riscontrare che nella maggior parte dei lavori (Richens e Lavigne, 1978; Patterson, 1988; McLaren e Green, 1985; Schultz e Bailey, 1978; MacArthur et al., 1982; Jeppensen 1987) ai fini di una più ampia conoscenza dell’impatto della caccia sugli ungulati è stata valutata la variazione della distanza di fuga in funzione di alcuni parametri come il sesso o la presenza di conspecifici, in altri casi, invece, sono state prese in considerazioni le variazioni stagionali degli home range (Root, 1998).
In questo studio sono state eseguite delle analisi, basate sul monitoraggio radiotelemetrico di un gruppo di 12 caprioli, che hanno interessato lo studio delle variazioni della dimensione degli home range, prendendo in considerazione fattori quali il sesso e le stagioni.
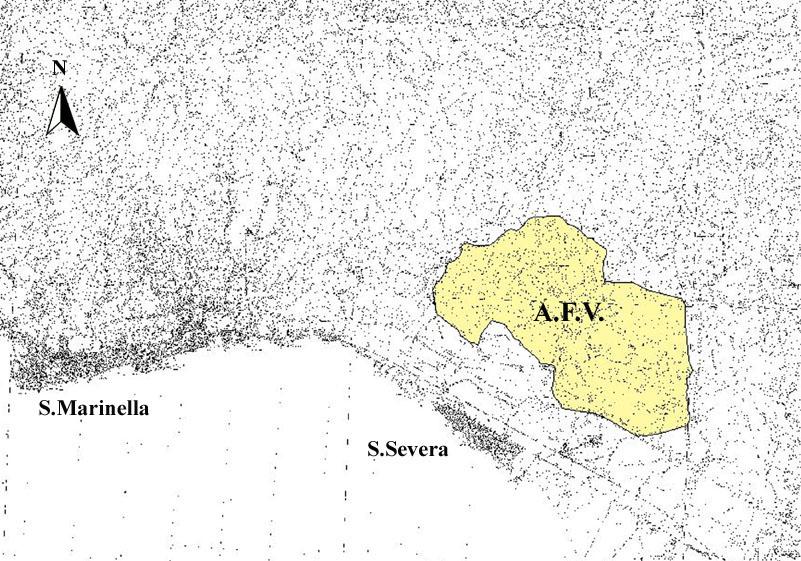
Lo studio è stato eseguito sui Monti della Tolfa (Provincia di Roma) e nello specifico nel territorio dell’Azienda Faunistica-Venatoria “Santa Severa” (fig 1.1) che è interamente compresa all’interno del comune di Tolfa e si estende su un territori di 1320 ha dei quali il 64% della superficie è ricoperta da boschi dove è praticata la ceduazione, il 32% da aree aperte, caratterizzate da pascoli pietrosi ed aridi o coltivati a graminacee mentre il restante 4% dalla macchia. Le quattro aree collinari più importanti sono quella di Monte Grande (309 m) e di Pian Sultano (263 m) separate dal fosso Eri, e quelle di Monte Fagiolano (161 m) e Monte del Mandriane (200 m).

Per quanto concerne la presenza di predatori per il capriolo, in generale si può affermare che essi sono assenti nonostante la presenza della volpe, del cinghiale e di qualche caso sporadico di randagismo, presenze che possono provocare predazione, soprattutto nei confronti dei piccoli caprioli.
La presenza antropica è limitata ad una piccola cava di caolino, da qualche escursionista in bicicletta o in moto, e, nel periodo invernale, dalla caccia e in più dalla gestione del bestiame, principalmente da bovini di razza maremmana, che in questa zona vengono allevati allo stato brado.
Nel periodo compreso tra il febbraio del 2001 e il febbraio del 2002 sono stati reintrodotti sui Monti della Tolfa, diciannove caprioli, di cui 8 maschi e 11 femmine, tutti originari della Tenuta Presidenziale di Castelporziano, poiché attualmente in essa risulta essere presente uno dei nuclei maggiori della sottospecie italica del capriolo, (C. c. italicus), insieme a piccole aree presenti nel Gargano e nel Pollino.
Come ultima cosa è stato applicato ad ogni animale il radiocollare provvisto di strie colorate di nastro isolante e di un numero di identificazione per il riconoscimento a distanza. A ogni animale è stato associato un identificativo numerico (ID) riportato su schede cartacee unitamente alla data, ora e luogo della cattura, condizioni di salute e frequenza del collare.
2. MATERIALI E METODI
I dati utilizzati per questo studio sono stati raccolti dal 2 luglio 2002 al 25 luglio 2004, per un totale di 753 giorni di campionamento e 4934 radiolocalizzazioni (fix) di cui solo 3791 sono stati utilizzati per questo studio. Le direzioni di provenienza del segnale sono state annotate su schede cartacee insieme alle stazioni di rilevamento e ad altre informazioni quali la data e l’ora, l’operatore, le condizioni meteo, l’attività dell’animale ed eventuali altre osservazioni.
I rilevamenti acquisiti con la tecnica del radio-tracking, restituiscono solamente una stima, più o meno accurata, della posizione dell’animale ad essa va necessariamente associata una misura dell’errore che si compie durante il rilevamento campionario.

E’ stato quindi introdotto il concetto di “area di confidenza” associata alla localizzazione stimata mediante triangolazione (Pedrotti e al., 1995). La forma e la dimensione dell’area di confidenza, e quindi la precisione di una localizzazione, dipendono dall’errore angolare (somma dell’errore di lettura ed errore del sistema), dalla distanza dell’animale (distanza tra radiocollare e ricevente) e dalla posizione relativa delle stazioni di rilevamento, che è ottimale quando gli angoli fra animale ed operatore sono di 90° (Saltz, 1994).
Una volta inseriti i dati in un database, per calcolare la localizzazione e l’area d’errore ad essa associata, è stato utilizzato il programma Locate II v.1.92 (Nams, 1990). Questo software utilizza per il calcolo della posizione il metodo MLE (maximum likelihood estimator, Lenth, 1981 in White e Garrott, 1990). Questo metodo produce una stima della posizione più probabile dell’animale e vi associa un’area di probabilità al 95% che, poiché si assume che l’errore di radiolocalizzazione sia una distribuzione normale bivariata, è di forma ellittica; che quindi descrive l’area dove si ha il 95% di probabilità di trovare l’animale al suo interno.

Bisogna considerare che la posizione delle stazioni di rilevamento influenza l’eccentricità e le dimensioni dell’ellisse di errore, in pratica la precisione della stima, ne consegue che le direzioni non possono essere prese da stazioni allineate, in modo che l’angolo formato dall’intersezione di due bearings non sia mai inferiore a 20° (Kenward, 1987; Millspaugh e Marzluff, 2001). Inoltre, poiché all’aumentare della distanza tra trasmittente e ricevente aumenta l’errore associato a ciascun bearing (Slade et al., 1965) si è cercato di utilizzare stazioni che fossero il più possibile vicino all’animale senza però interferire con l’attività di quest’ultimo.
Un’ulteriore fonte di errore nella stima della posizione dell’animale risulta dal fatto che i bearings vengono raccolti sequenzialmente e non simultaneamente. Ciò implica che durante la triangolazione l’animale potrebbe muoversi producendo così un errore nella stima della sua posizione (Cochran, 1980; White e Garrott, 1990). Si è cercato, perciò, di far passare il minor tempo possibile tra il primo e l’ultimo bearings, in genere 10 – 15 minuti.

Per rendere più precisa la stima della posizione dell’animale e dell’ellisse di errore, per ogni operatore e per ciascuno dei quattro semestri in cui è stato suddiviso il periodo di studio, sempre in relazione al singolo animale, è stata calcolata la deviazione standard (SD) media nella rilevazione del bearing; si è riscontrata una differenza di precisione tra i diversi operatori. Tutte le ellisse di errore sono state quindi ricalcolate dal programma immettendo il valore opportuno di SD, per ottenere stime più precise dell’area di errore.
3. ANALISI DEI DATI
3.1 Home-range
I dati radiotelemetrici , esclusi quelli delle continuate, sono stati utilizzati per il calcolo degli home range (HR), o area vitale con il quale, secondo Burt (1943), si intende “l’area utilizzata da un individuo durante le sue normali attività di ricerca del cibo, accoppiamento e allevamento della prole”, ovviamente nel determinare l’home range di un animale svolge un ruolo fondamentale sia la scala temporale sulla quale esso viene calcolato, sia il metodo di calcolo utilizzato per la sua stima.
Anche lo stato dell’individuo considerato (sesso, età, classe,ruolo sociale, stato riproduttivo, ecc.) è un parametro importante per la definizione di un home range (Harris e al., 1990) (White & Garrot, 1990).
I metodi per calcolare l’home range sono tre: in poligonali, grid-cell e probabilistici. I primi costruiscono l’HR connettendo le localizzazioni periferiche di un animale, secondo regole precise, per formare poligoni concavi o convessi; i metodi “grid-cell” usano una griglia per discretizzare l’area studiata in maniera più o meno fine; infine nei metodi probabilistici viene determinata la probabilità di incontrare l’individuo in ciascun punto dello spazio, associandovi un intervallo di confidenza; in questo caso il modello di uso dello spazio è conforme a priori ad una particolare distribuzione di probabilità (Pedrotti e al., 1995). Con i metodi probabilistici è possibile identificare, all’interno dell’home-range, l’esistenza di zone più intensamente utilizzate, in quanto sedi di funzioni biologiche (siti di alimentazione, aree del parto): le core area (Harris et al., 1990) essa è la parte dell’home range dove vi è una maggiore intensità di localizzazioni e la sua determinazione nasce dalla considerazione che ogni animale usa il proprio areale con diversa intensità..
Ognuno di questi 3 metodi analitici ha i suoi assunti, i suoi vantaggi ed i suoi difetti ed ovviamente nessuna tecnica è di per sé perfetta.
Il Minimo Poligono Convesso (MCP) (Mohr, 1947) è il più antico, immediato e comune metodo di stima dell’home range (White e Garrott, 1990). Esso calcola l’area più piccola contenente tutte le localizzazioni; il poligono che né deriva è convesso giacché tutti gli angoli interni sono inferiori a 180°. Tale metodo non fornisce, quindi, informazioni sui centri di attività, valgono a dire sulle zone maggiormente frequentate dagli animali, ma considera ogni sezione dell’home range usata con uguale frequenza (White e Garrott, 1990); può, inoltre, includere zone mai frequentate. Il vantaggio è che non necessita di alcuna assunzione sulla distribuzione dei dati; è inoltre possibile, con tale metodo, confrontare i risultati ottenuti nei diversi studi che ne hanno fatto uso. In questo studio si è scelto di eliminare il 5% delle localizzazioni dal calcolo dell’MCP, stimando le localizzazioni estreme con il metodo della media armonica (Dixon & Chapman, 1980).
Il metodo della media armonica è un metodo non parametrico e, come tale, non comporta l’assunzione a priori della forma della UD come circolare o bivariata, come avviene per i metodi parametrici (Pedrotti et al., 1995), ma stima questa funzione dai dati.
Questo metodo utilizza la formula della media armonica
HMj=1/(1/P)[Σp(1/rjx)]
(dove P è il numero di osservazioni, j è ogni punto di una griglia sovrimposta e r il raggio tra l’osservazione x e il punto j) come misura dell’intensità di utilizzo dei vari punti dello spazio. Per ogni nodo j della griglia viene calcolato il valore della media armonica (HM) e tutti i punti con HM uguale vengono uniti costruendo delle isoplete che racchiudono percentuali di localizzazioni crescenti, a partire dalle zone più intensamente utilizzate. Il limite di questo metodo è quello di essere fortemente influenzato dalla posizione e dalla dimensione della griglia sovrimposta (Dixon & Chapman, 1980).
In questo modo, le localizzazioni che presentavano valori di media armonica più elevati sono state via eliminate, ricalcolandone ogni volta il valore fino ad avere un campione che contenesse solo il 95% dei fix originari (White & Garrot, 1990). Questa operazione è stata svolta usando la funzione Outlier Removal, dall’estensione Animal Movement Analysis (USGS, 1998) del programma “ArcView GIS (Geographic Information System)” (ESRI, 1998). Per il calcolo del 95% MCP si è utilizzata la funzione Minimum Convex Polygon Home Range della stessa estensione del software.
Tra i metodi probabilistici si è scelto quello di Kernel (KHR)(Worton, 1987) , perché considerato il più efficace nel calcolo di dimensione e forma dell’home-range.
Con questo metodo, che permette di stimare la distribuzione di utilizzo (UD), cioè la distribuzione di frequenza delle localizzazioni nel tempo e nello spazio (Van Winkle, 1975), si calcola la funzione di probabilità a partire da una sovrapposizione di griglia ad un set di localizzazioni (Pedrotti e al., 1995). Un aspetto importante, quando si procede al calcolo dell’home range tramite il metodo di kernel, è il parametro h di smoothing (o di addolcimento); questo parametro è riconosciuto essere il componente critico di questo metodo. A tal riguardo esistono due diversi tipi di calcolo dello stimatore di kernel: nel kernel fisso lo stesso parametro di smoothing viene usato per ogni localizzazione in ogni punto della distribuzione; questo metodo generalmente produce stime più accurate e precise soprattutto lungo i contorni esterni dell’home range, mentre perde precisione nelle aree interne ad alta densità di localizzazioni (Seaman e al., 1999). Nel kernel adattativo invece viene selezionato un parametro di smoothing per ogni localizzazione, maggiore nelle aree con poche osservazioni e minore in aree con molte. Questo metodo produce una moderata accuratezza nella stima con un’elevata precisione nelle aree interne ad alta densità di localizzazioni (Seaman e al., 1999).
Nel presente studio si è utilizzato il Kernel adattativo ed il parametro di smoothing è stato scelto ad hoc mediante il metodo Least Square Cross Validation (LSCV) (Silvermann, 1986). Usualmente la stima della distribuzione d’uso viene calcolata su una percentuale fissa di localizzazioni che, nel nostro caso, corrisponde al 95%, 70% e 20% per gli home range ed al 50% per le core area (Harris et al., 1990; Pedrotti et al., 1995). La core-area è la parte dell’home range dove vi è una maggiore intensità di localizzazioni e la sua determinazione nasce dalla considerazione che ogni animale usa il proprio areale con diversa intensità.
In questo lavoro si è usato il metodo del kernel fisso al 95%, 70% , 50%e 20%; per effettuare i calcoli, il database delle localizzazioni di tutti gli animali è stato importato nel programma Arc View 3.2a che, tramite l’estensione Animal Movement 2.04b (Esri, 2000), permette di calcolare gli home range.
Per le analisi degli Home range, una volta accertata la significatività di ogni variabile, tramite il test di normalità di Shapiro, si è utilizzato il Modello Lineare Generalizzato (GLM) per testare gli effetti delle variabili sesso e stagioni.
Verificata la significatività del modello generale (Model) sono state confrontate le medie con il test di Bonferroni.
3.2 Scala temporale
Nel presente studio, tutte le analisi sono state effettuate sia scala annuale, considerando quindi i due anni di ricerca (Tab. 4.3), sia su scala stagionale, dividendo ogni anno in quattro distinte stagioni di tre mesi ciascuna, per un totale di sette stagioni (Tab. 4.4).
|
Periodo |
Anno |
|
02 luglio 2002 – 01 luglio 2003 |
1 |
|
02 luglio 2003 – 25 luglio 2004 |
2 |
Tabella 3.1 Definizioni degli anni.
|
Periodo |
Identificativo |
|
01 novembre – 31 gennaio |
1 |
|
01 febbraio – 30 aprile |
2 |
|
01 maggio – 31 luglio |
3 |
|
01 agosto – 31 ottobre |
4 |
Tabella 3.2 Definizione delle stagioni
Questa scelta è stata effettuata relazionando i comportamenti ed i ritmi di attività del capriolo con i periodi di caccia al cinghiale.
4. RISULTATI
4.1 Descrizione del campione
Complessivamente il campione analizzato è composto di 12, di cui 5 maschi e 7 femmine radiolocalizzati a partire dal 2 luglio 2002. Nel periodo tra il 29 luglio 2003 e 31 dicembre 2003, il numero di animali è sceso a 7 i quanto sono andati a esaurimento le batterie dei collari di 4 animali, T05, T06, T07 e il T12. Il capriolo T03 è stato vittima, invece, di un atto di bracconaggio nei pressi di Monte Lungo, dove è stato rinvenuto il solo collare.
Il monitoraggio radiotelemetrico su gli individui rimasti si è protratto fino al 25 luglio 2004 con un totale di 4934 radiolocalizzazioni suddivisi in fix singoli giornalieri e in fix rilevati durante le continuate.. Dopo le operazioni preliminari, che hanno previsto sia l’esclusione dei fix con un’area superiore ai 10 ha sia l’esclusione dall’analisi dei fix di quegli individui che possedevano un numero di radiolocalizzazioni troppo piccolo per ogni singola stagione; il set di dati è risultato composto da 3791 localizzazioni. (Harris et al., 1990; Ronchi, 2000).
Nella tabella 4.1 vengono riportato il numero delle localizzazioni che sono state utilizzate per il calcolo degli Home range annuali e stagionali.
|
|
1 Anno |
2 Anno |
|||||
|
ID |
1 Stg |
2 Stg |
3 Stg |
4 Stg |
1 Stg |
2 Stg |
3 Stg |
|
T03 |
70 |
34 |
36 |
35 |
4 |
———- |
———- |
|
T05 |
69 |
34 |
36 |
35 |
28 |
———- |
———- |
|
T06 |
69 |
34 |
36 |
33 |
8 |
———- |
———- |
|
T07 |
67 |
34 |
36 |
35 |
4 |
———- |
———- |
|
T09 |
69 |
34 |
36 |
35 |
31 |
30 |
30 |
|
T11 |
70 |
34 |
35 |
35 |
31 |
30 |
30 |
|
T12 |
69 |
34 |
36 |
———- |
———- |
———- |
———- |
|
T13 |
68 |
34 |
36 |
32 |
31 |
30 |
29 |
|
T14 |
68 |
34 |
34 |
35 |
31 |
30 |
30 |
|
T15 |
69 |
34 |
35 |
34 |
31 |
30 |
30 |
|
T16 |
69 |
34 |
36 |
33 |
29 |
30 |
30 |
|
T19 |
69 |
34 |
36 |
34 |
31 |
37 |
29 |
Tabella 4.1 Numero di localizzazioni per animale, per stagione e per anno
4.2 Analisi dell’Home range
Gli home range annuali sono stati calcolati con il metodo Kernel al 20%,50%,70% e 95%, . Nelle tabelle sottostanti sono riportati i valori dei vari home range (Kernel al 20%,50%,70% e 95%) in funzione del periodo complessivo di studio, delle stagioni e del sesso. Si è voluto anche aggiungere, ai fini di una più completa chiarezza, il numero di campione testato e il valore massimo e minimo che si è riscontrato per una determinata categoria.
4.2.1 Analisi preliminari degli Home range annuali
Prima dell’analisi vera e propria, , si sono voluti presentare alcuni dei dati utili per la comprensione delle dimensioni, in ettari, degli andamenti annuali degli home range., sia per tutti gli animali che per i due sessi distinti.
|
Home range |
Anno |
Numero del campione |
media |
Valore minimo e massimo |
deviazione standard |
|
KHR_20 |
1 |
12 |
2.51 |
0.69 / 5.19 |
1.47 |
|
2 |
8 |
5.32 |
0.83 / 19.78 |
5.75 |
|
|
KHR_50 |
1 |
12 |
11.55 |
4.95 / 19.54 |
5.18 |
|
2 |
8 |
23.003 |
3.51 / 80.01 |
23.25 |
|
|
KHR_70 |
1 |
12 |
25.15 |
9.15 / 48.10 |
12.59 |
|
2 |
8 |
43.86 |
7.84 / 155.28 |
42.65 |
|
|
KHR_95 |
1 |
12 |
90.41 |
25.64 / 210.85 |
53.26 |
|
2 |
8 |
105.98 |
22.25 / 401.68 |
104.12 |
Tabella 4.2 Ampiezza in ettari degli home range annuali calcolati con il metodo Kernel
Tutti gli home range annuali calcolati tramite il metodo di Kernel, al 20%,50%,70% e 95%, considerando i due sessi separatamente, mettono in evidenza l’assenza di significatività tra i maschi e le femmine.
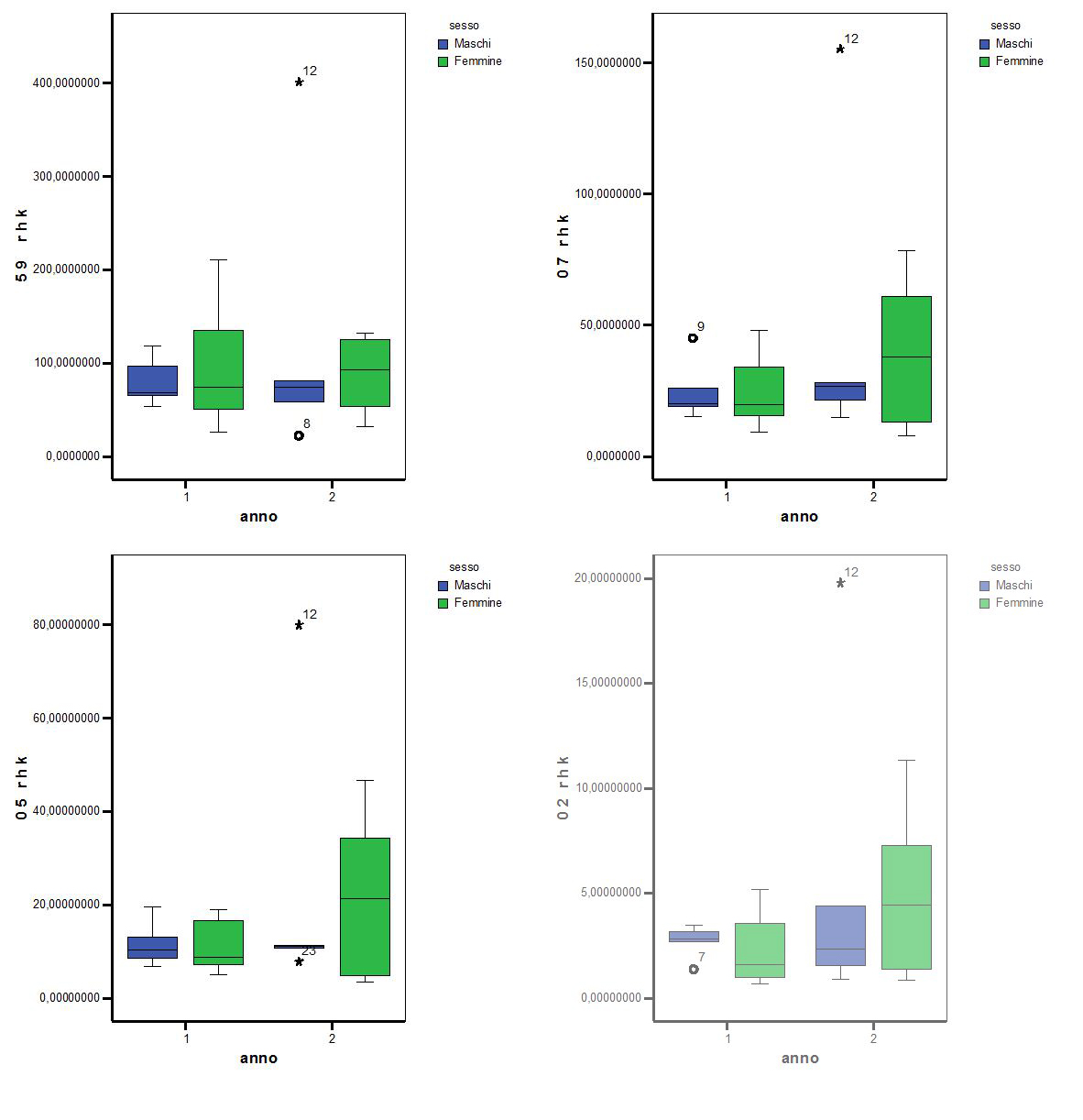
(anno 1 e 2) considerando i sessi separati
4.2.2 Analisi preliminari degli Home range stagionali
Anche per l’analisi degli home range stagionali si sono voluti presentare alcuni dei dati utili alla comprensione delle dimensioni, in ettari, degli andamenti degli home range., sia per tutti gli animali che per i due sessi distinti.
|
Home range |
stg |
Numero del campione |
media |
Valore minimo e massimo |
deviazione standard |
|
KHR_20 |
1 |
20 |
4.77 |
0.6 / 19.59 |
4.73 |
|
2 |
19 |
2.87 |
0.34 / 10.72 |
2.72 |
|
|
3 |
19 |
0.94 |
0.37 / 2.18 |
0.48 |
|
|
4 |
11 |
1.6 |
0.25 / 7.96 |
2.15 |
|
|
KHR_50 |
1 |
20 |
20.13 |
3.34 / 81.28 |
20.09 |
|
2 |
19 |
11.08 |
2.53 / 33.58 |
8.33 |
|
|
3 |
19 |
4.73 |
1.65 / 9.36 |
2.22 |
|
|
4 |
11 |
5.6 |
1.95 / 19.91 |
4.88 |
|
|
KHR_70 |
1 |
20 |
40.32 |
8.88 / 155.55 |
38.25 |
|
2 |
19 |
22.55 |
6.22 / 57.60 |
14.15 |
|
|
3 |
19 |
10.85 |
4.01 / 177.93 |
4.42 |
|
|
4 |
11 |
12.83 |
7.01 / 37.86 |
8.69 |
|
|
KHR_95 |
1 |
20 |
107.53 |
28.04 / 286.62 |
71.74 |
|
2 |
19 |
68.17 |
20.72 / 187.45 |
39.93 |
|
|
3 |
19 |
30 |
12.39 / 69.04 |
15.52 |
|
|
4 |
11 |
37.54 |
16.20 / 80.48 |
16.72 |
Tabella 4.3 Ampiezza in ettari degli home range stagionali calcolati con il metodo Kernel
Tutti gli home range stagionali delle femmine, calcolati tramite il metodo di Kernel, al 20%,50%,70% e 95%, risultano essere,sia per il primo anno che per il secondo anno, più grandi rispetto a quelli dei maschi.
Analizzando nello specifico gli home range, sia delle femmine che dei maschi, si può dedurre che la stagione del periodo invernale risulta sempre quella con un home range più grande rispetto alle altre stagioni.
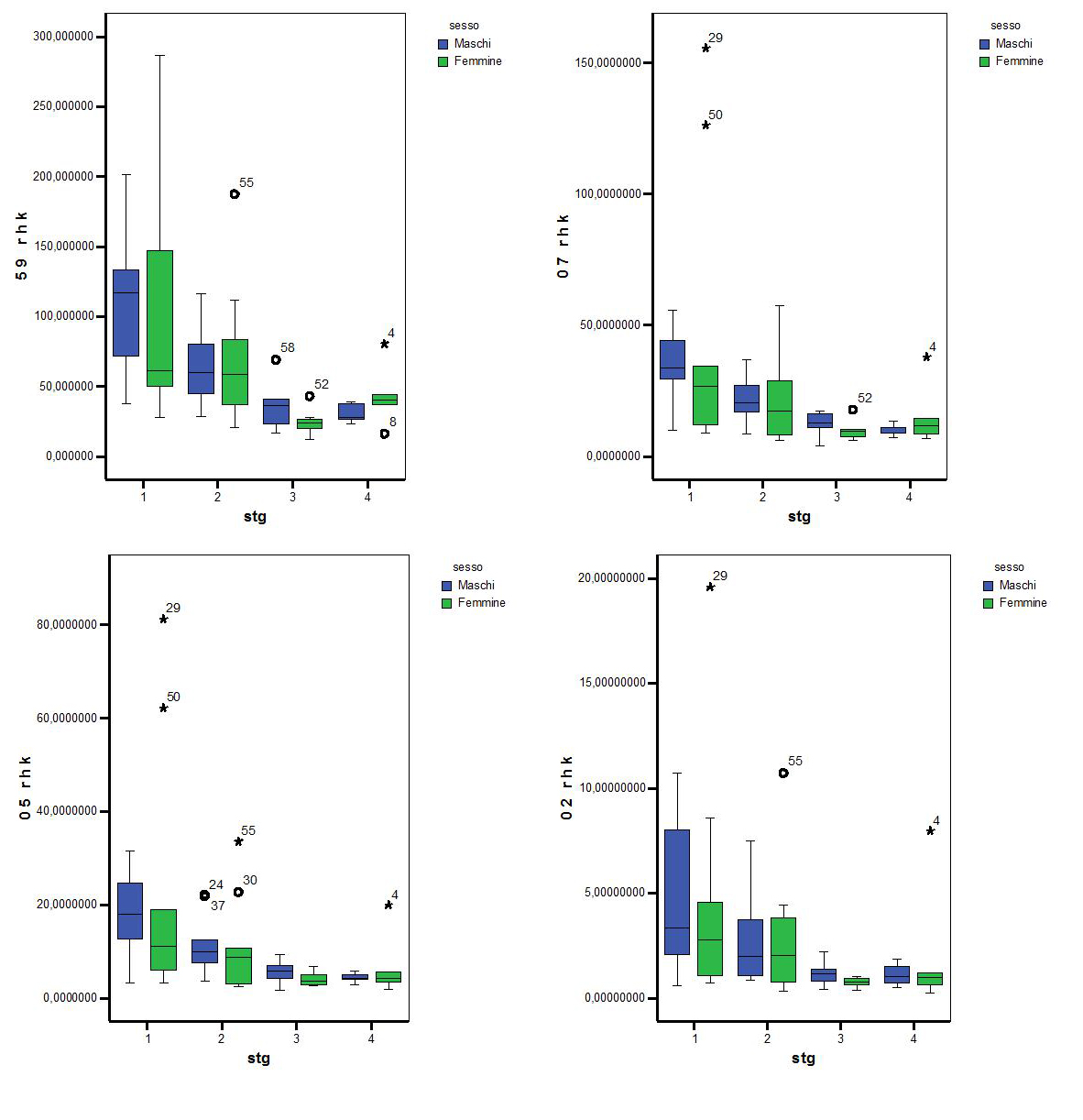
stagionali ( stg1, stg2 , stg3, stg4 ) considerando i sessi separati
4.2.3 Periodo totale di studio
Lo studio dell’home range è stato eseguito con il test del GLM in SAS considerando sia il periodo di studio nella sua totalità che il primo e secondo anno separatamente.
Tutti e tre i periodi sono stati analizzati in funzione delle variabili sesso, stagione e dalla interazione tra sesso e stagione
Il modello generale per quanto riguarda l’intero periodo di studio risulta significativo. Delle variabili analizzate solamente quella riguardante la stagione è risultata essere significativa mentre sia per il sesso che per l’interazione tra la stagione ed il sesso i valori non hanno dato risultati inferiori alla soglia dello 0,05 risultando, quindi, non significativi.
|
Periodo totale di studio |
|||
Kernel 20% |
|||
|
df |
F |
P |
|
|
Model |
7 |
3.04 |
0.0092 |
|
Sesso |
1 |
0.65 |
0.4254 |
|
Stg |
3 |
6.53 |
0.0008 |
|
Sesso*Stg |
3 |
0.1 |
0.9585 |
Tabella 4.4 Risultati del test GLM sul campione totale degli home range con Kernel 20%
Eseguendo il test di Bonferroni sugli Home Range al Kernel al 20% e sul periodo totale di studio, risultano significativamente diverse alcune stagioni. In particolare è la stagione 1 (media 3.123 ettari) che risulta differente sia dalla stagione 3 (medi 0.868 ettari) che dalla stagione 4 (media 1.042 ettari) .
Periodo totale di studio |
|||
Kernel 50% |
|||
|
df |
F |
P |
|
|
Model |
7 |
3.66 |
0.0027 |
|
Sesso |
1 |
0.33 |
0.5693 |
|
Stg |
3 |
8.09 |
0.0002 |
|
Sesso*Stg |
3 |
0.15 |
0.9288 |
Tabella 4.5 Risultati del test GLM sul campione totale degli home range con Kernel 50%
Eseguendo il test di Bonferroni sugli Home Range al Kernel al 50% e sul periodo totale di studio, risultano significativamente diverse alcune stagioni.
In particolare è la stagione 1 (media 13.8985 ettari) che risulta differente sia dalla stagione 3 (media 4.4315 ettari) che dalla stagione 4 (media 4.586 ettari).
|
Periodo totale di studio |
|||
Kernel 70% |
|||
|
df |
F |
P |
|
|
Model |
7 |
3.99 |
0.0014 |
|
Sesso |
1 |
0.01 |
0.9090 |
|
Stg |
3 |
8.91 |
< .0001 |
|
Sesso*Stg |
3 |
0.27 |
0.8491 |
Tabella 4.6 Risultati del test GLM sul campione totale degli home range con Kernel 70%
Eseguendo il test di Bonferroni sugli Home Range al Kernel al 70% e sul periodo totale di studio, risultano significativamente diverse alcune stagioni. In particolare è la stagione 1 (media 29.73 ettari) che risulta differente sia dalla stagione 3 (media 10.07 ettari) che dalla stagione 4 (media 11.62 ettari).
|
Periodo totale di studio |
|||
Kernel 95% |
|||
|
df |
F |
P |
|
|
Model |
7 |
5.79 |
< .0001 |
|
Sesso |
1 |
0.32 |
0.5726 |
|
Stg |
3 |
12.30 |
< .0001 |
|
Sesso*Stg |
3 |
0.64 |
0.5956 |
Tabella 4.7 Risultati del test GLM sul campione totale degli home range con Kernel 95%
Eseguendo il test di Bonferroni sugli Home Range al Kernel al 95% e sul periodo totale di studio, risultano significativamente diverse alcune stagioni. In particolare è la stagione 1 (media 87.32 ettari) che risulta differente sia dalla stagione 3 (media 27.65 ettari) che dalla stagione 4 (media 34.11 ettari) ,mentre la stagione 2 (media 58.53 ettari) risulta essere differente dalla stagione 3 (media 27.65 ettari).
4.2.4 Primo e secondo anno
Anno 1 |
Anno 2 |
|||||
Kernel 20% |
||||||
|
F |
P |
df |
F |
P |
df |
|
|
Model |
3.80 |
0.0031 |
7 |
0.73 |
0.5584 |
3 |
|
Sesso |
0.77 |
0.3849 |
1 |
0.13 |
0.7281 |
1 |
|
Stg |
8.27 |
0.0002 |
3 |
1.33 |
0.2763 |
1 |
|
Sesso*Stg |
0.13 |
0.9419 |
3 |
1.02 |
0.3364 |
1 |
Tabella 4.8 Risultati del test GLM sul campione, diviso tra primo e
secondo anno, degli homerange con Kernel 20%
Eseguendo il test di Bonferroni sugli Home Range al Kernel al 20% e sul primo anno di studio, risultano significativamente diverse alcune stagioni. In particolare è la stagione 1 (media 4.28 ettari) che risulta differente dalla stagione 2 (media 1.58 ettari), dalla stagione 3 (media 0.8686 ettari) e dalla stagione 4 (media 1.042 ettari).
Per il secondo anno di studio non è stato eseguito il test di Bonferroni perché il modello generale non risulta significativo.
Anno 1 |
Anno 2 |
|||||
Kernel 50% |
||||||
|
F |
P |
df |
F |
P |
df |
|
|
Model |
4.68 |
0.0007 |
7 |
0.97 |
0.4429 |
3 |
|
Sesso |
0.29 |
0.5932 |
1 |
0.35 |
0.5664 |
1 |
|
Stg |
10.36 |
< .0001 |
3 |
1.79 |
0.2103 |
1 |
|
Sesso*Stg |
0.19 |
0.9057 |
3 |
1.13 |
0.3120 |
1 |
Tabella 4.9 Risultati del test GLM sul campione, diviso tra primo e
secondo anno, degli homerange con Kernel 50%
Eseguendo il test di Bonferroni sugli Home Range al Kernel al 50% e sul primo anno di studio, risultano significativamente diverse alcune stagioni. In particolare è la stagione 1 (media 18.53 ettari) che risulta differente dalla stagione 2 (media 6.81 ettari), dalla stagione 3 (media 4.304 ettari) e dalla stagione 4 (media 4.58 ettari).
Per il secondo anno di studio non è stato eseguito il test di Bonferroni perché il modello generale non risulta significativo.
Anno 1 |
Anno 2 |
|||||
Kernel 70% |
||||||
|
F |
P |
df |
F |
P |
df |
|
|
Model |
4.62 |
0.0008 |
7 |
0.73 |
0.5559 |
3 |
|
Sesso |
0.06 |
0.8156 |
1 |
0.01 |
0.9177 |
1 |
|
Stg |
10.25 |
< .0001 |
3 |
1.17 |
0.3047 |
1 |
|
Sesso*Stg |
0.31 |
0.8152 |
3 |
1.02 |
0.3373 |
1 |
Tabella 4.10 Risultati del test GLM sul campione, diviso tra primo e
secondo anno, degli homerange con Kernel 70%
Eseguendo il test di Bonferroni sugli Home Range al Kernel al 70% e sul primo anno di studio, risultano significativamente diverse alcune stagioni. In particolare è la stagione 1 (media 37.69 ettari) che risulta differente dalla stagione 2 (media 15.32 ettari), dalla stagione 3 (media 37.69 ettari) e dalla stagione 4 (media 11.13 ettari).
Per il secondo anno di studio non è stato eseguito il test di Bonferroni perché il Modello generale non risulta significativo.
Anno 1 |
Anno 2 |
|||||
Kernel 95% |
||||||
|
F |
P |
df |
F |
P |
df |
|
|
Model |
6.43 |
< .0001 |
7 |
0.76 |
0.5393 |
3 |
|
Sesso |
0.39 |
0.5348 |
1 |
0.08 |
0.7855 |
1 |
|
Stg |
13.93 |
< .0001 |
3 |
1.40 |
0.2649 |
1 |
|
Sesso*Stg |
0.68 |
0.5710 |
3 |
1.14 |
0.3117 |
1 |
Tabella 4.11 Risultati del test GLM sul campione, diviso tra primo e
secondo anno, degli homerange con Kernel 95%
Eseguendo il test di Bonferroni sugli Home Range al Kernel al 95% e sul primo anno di studio, risultano significativamente diverse alcune stagioni. In particolare è la stagione 1 (media 108.81 ettari) che risulta differente dalla stagione 2 (media 47.92 ettari), dalla stagione 3 (media 27.78 ettari) e dalla stagione 4 (media 34.24 ettari).
5. DISCUSSIONE
Nel quadro dei risultati ottenuti nel presente studio si evidenzia una differenza significativa tra le quattro diverse stagioni ed in particolare risulta che nella prima stagione, quella invernale che comprende tutto il periodo di caccia, i caprioli hanno in media dimensioni più grandi dell’home range.
I risultati ottenuti da altri studi, effettuati sulla specie sia in ambienti di foresta che in ambienti di campo, (Cederlund, 1983; Dzieciolowski e al., 1990; Koubek 1995, Tufto e al., 1996; Jewell 1966; Sanderson 1966; Clutton-Brock & Harvey 1978 ) mettono in risalto che uno dei principali fattori a determinare un aumento o una diminuzione dell’area dell’home range è la disponibilità ed accessibilità di alimento.
In particolare, da i risultati ottenuti da Cederlund (1983), nel nord Europa, in ambiente di foresta, gli home range della stagione primaverile-estiva risultano più grandi di quelli che invece si ottengono nella stagione autunnale-invernale e questa differenza viene imputata sia alla diversa disponibilità di cibo sia alle diverse condizioni climatiche, variabili che ovviamente cambiano nei diversi periodi dell’anno.
Negli studi di Dzieciolowski (1990), Jewell ( 1966), Sanderson e Clutton-Brock & Harvey (1978), le oscillazione delle dimensioni degli home range vengono imputate sia alla disponibilità di cibo sia anche alle variazioni nella densità degli animali. In particolare, nello studio di Dzieciolowski, nel periodo che comprende i mesi tra novembre e marzo, a causa della più alta densità degli animali, gli home range hanno una estensione maggiore, rispetto a quelli registrati nel periodo di tempo compreso tra aprile e ottobre.
Al contrario, nello studio di Jeppesen (1987), sul cervo, l’aumento delle dimensioni degli home range riscontrato nella stagione invernale viene imputato all’aumento del disturbo antropico.
I risultati da noi ottenuti sono confrontabili con quelli ottenuti in questo ultimo studio. L’ipotesi più plausibile, riguardo all’aumento di dimensioni degli home range nella stagione invernale, può essere imputato alla presenza della caccia. In particolare questo continuo e assiduo disturbo, che viene effettuato per cinque giorni alla settimana, per tutta la durata della stagione, può provocare un aumento a partire dalla propria core area, con conseguente ingrandimento dell’area occupata dal capriolo.
Considerando separatamente i maschi e le femmine, non risultano significative differenze tra le aree calcolate a diverse percentuali di utilizzo degli home range sia a livello annuale che a livello stagionale. Questo conferma i risultati presentati sia da Koubek (1995), da Catt (1986) e da Dzieciolowski et al. (1990) mentre è in contrasto sia con quelli di Cederlund (1983) e Mysterud (1998),dove i maschi presentano home range più grandi rispetto a quelli delle femmine a causa della diversa richiesta di cibo, sia con lo studio di Marchal (e al., 1998) dove sono le femmine quelle con home range più grandi rispetto ai quelli dei maschi.
Dallo studio di Jeppensen (1987) i disturbi possono essere divisi in predittibili e non-predittibili, e nei primi rientrano i casi di disturbo dato dal traffico mentre nei secondi quei disturbi apportati dal turismo e dalla caccia.
Gli unici casi nei quali sembra ci sia stata abituazione al disturbo riguardano tutte situazioni che non presentavano mai attività venatoria ma disturbi legati o al traffico o al turismo (Schultz e Bailey, 1978; MacArthur et al., 1982; Patterson, 1988). Studi eseguiti in merito alla abituazione al disturbo non hanno riguardato solo i caprioli ma anche altre specie di ungulati. Sono stati fatti lavori sul cervo mulo (Richens e Lavigne, 1978), sul camoscio (Patterson, 1988 ) e sul bue muschiato (McLaren e Green, 1985) e in nessuno di questi studi è stato possibile parlare di assuefazione al disturbo antropico.
Per quanto concerne, in particolare, il disturbo apportato dalla caccia nei riguardi del capriolo, risulta che questi animali, tra i più riservati e schivi della famiglia dei Cervidi, subiscono sempre un forte stress (Jeppensen, 1987). Questo stress può manifestarsi con l’allontanamento dal proprio home range, anche per più giorni, con una minore stabilità all’interno del proprio territorio, con una più elevata distanza di fuga favorita dalla presenza di cani e di cacciatori nel momento in cui si dispongono ed successivamente danno iniziano alla battuta di caccia. In diversi studi (Jeppensen, 1987 (3); MacArthur et al., 1982; Root et al., 1988; Sonetti e Galimberti, 1991; Jeppensen, 1987 (2); de Boer et al., 2003) è stato trovato che nelle femmine si osserva una vigilanza sopra il normale nei luoghi dove viene praticata la caccia e, più in generale, il comportamento dei singoli individui presenta una maggiore riservatezza sempre dove la caccia è costantemente praticata.
Questo lavoro ha permesso di approfondire le conoscenze, ad oggi ancora scarse, del capriolo italico ed in particolare analizzando una popolazione reintrodotta con l’impatto della caccia al cinghiale secondo lo studio degli home range.
I risultati ottenuti evidenziano che gli animali sono sottoposti ad uno stress particolarmente significativo quando viene praticata la caccia al cinghiale e che l’impatto provocato dall’uomo non è circoscritto solamente al tempo della cacciata, ma che persiste anche nei periodi seguenti.
La prova, di quanto affermato sopra, è data dal fatto che gli animali, durante il periodo di caccia presentano sia degli home range notevolmente più grandi.
Alla luce di quanto detto è quindi di fondamentale importanza, ai fini di una migliore gestione e protezione della specie, valutare sempre che nelle zone dove viene praticata la caccia, gli animali vivono in una condizione di elevato stress per un certo periodo di tempo. Questa situazione, come abbiamo visto, influenza il comportamento abituale degli individui e di ciò è opportuno tenerne conto nell’intraprendere un progetto di reintroduzione.
BIBLIOGRAFIA
- AANES A.; ANDERSEN R. (1996). The effect of sex, time of birth, and habitat on the vulnerability of roe deer fawns to red fox predation. Can. J. Zool. 74 : 1857-1865.
- AANES R.; LINNELL J.D.C.; PERZANOWSKI K.; KARLSEN J.; ODDEN J. (1998). Roe deer as a prey. The European roe deer: the biology of success (eds Andersen R., Duncan P., Linnell J.D.C.). Scandinavian University Press. Pp. 377.
- AEBISCHER N.J., ROBERTSON P.A., 1992. Pratical aspects of compositional analysis as applied to pheasant habitat utilization. In: Wildlife Telemetry: Remote monitoring and Tracking of Animals. Priede & Swift : 285-293.
- AEBISCHER N.J., ROBERTSON P., KENWARD R.E.,1993. Compositional analysis of habitat use from animal radio-tracking data. Ecology, 74(5): 1313-1325.
- Andersen R., Duncan P., Linnel J., 1998. The European Roe Deer: the biology of success. Scandinavian Universoty Press. Oslo. 376 pp.
- AULAK W., BABINSKA-WERKA J., 1990. Preference of different habitats and age classes of forest by roe deer. Acta Theriologica 35(3-4): 289-298.
- BIDEAU E., GERARD J.F., VINCENT J.P., MAUBLANC M.L., 1993. Effects of age and sex on space occupation by european roe deer. J. Mamm., 74(3):745-751.
- BIDEAU E., VINCENT J.P., MAUBLANC M.L., GONZALEZ R., 1987. Dispersion chez le jeune chevreuil (Capreolus capreolus L.) : étude sur une population en milieu forestier. Acta Oecologica- Oecol. Applic., 8(2):135-148.
- BIDEAU E., VINCENT J.P., QUERE J.P., ANGIBAULT J.M., 1983. Occupation de l’espace chez le chevreuil (Capreolus capreolus L., 1789). Acta Oecologica-Oecol. Applic., 4(2):163-184.
- BRUNO S., 1977.Gli anfibi e i rettili della Tolfa (Antiappennino laziale). In AA.VV., Problemi attuali di scienza e di cultura. Ricerche ecologiche , floristiche e faunistiche nel comprensorio Tolfetano- Cerite- Manziate. Accademia Nazionale dei Lincei, 227: 89-124.
- BRUNO E., CORSINI M.T., DRAGHI A., MORIMANDO F., 1998. Relazioni tra habitat, densità e caratteristiche biometriche del capriolo in alcune aree della provincia di Grosseto. II Congresso Italiano di Theriologia “I mammiferi in Italia:status, tendenze e implicazioni gestionali”. Varese 28-30 Ottobre: 112.
- BURT W.H., 1943. Territoriality and home range concepts as applied to mammals. Journal of Mammalogy 24: 346-352.
- CALANGE C, MAILLARD D., INVRNIA e GAUDIN, 2005. Reinttroduction or roe deer Capreolus capreolus into a Mediterraneam habitat: female mortality and dispersion. Wild. Biol. 11: 153-161.
- CARNEVALI L., 1998. Uso e selezione dell’habitat di femmine di capriolo (Capreolus capreolus) nell’Appennino settentrionale. Tesi di Laurea, Università degli studi di Roma “La Sapienza”.
- CaTT D.C. , and STAINES B. W., 1986. HOME RANGE USE HABITAT SELECTION BY Red deer (Cervus alaphus) in a Sitka spruce plantation as determined by radio-tracking. J.Zool.,Lond 211,681-693.
- Cederlund G., Linstrom E., 1983. Effects of severe winters and fox predation on roe deer mortality. Acta Theriologica, 28(7):129-145.
- CHIAVETTA M., 1977. Ricerche sugli uccelli rapaci del comprensorio Tolfetano-Cerite. In AA.VV., Problemi attuali di scienza e di cultura. Ricerche ecologiche , floristiche e faunistiche nel comprensorio Tolfetano- Cerite- Manziate. Accademia Nazionale dei Lincei, 227: 177- 190.
- CIANFANELLI L.; ROVEDA D.; CHIODINI R.; FRUGIS S.; PERCO F.; CRAVIN A.; PETTITI L. (1997). Liberazione da recinto di Capriolo (Capreolus capreolus) per la reintroduzione nel Parco lombardo della Valle del Ticino.Suppl. Ric. Biol. Selvaggina XXVII: 453-456.
- CIBIEN C., BIDEAU E., BOISAUBERT B., BIRAN H., ANGIBAULT J.M., 1995. Seasonal diet and habitat use in field roe deer (Capreolus capreolus) in the Picardie region. Gibier Faune Sauvage, Game Wildlife Vol 12: 37-49.
- Cibien C., Bideau E., Boisaubert B., Maublanc M.L., 1989. Influence of habitat characteristics on winter social organisation in field roe deer. Acta Theriologica, 34: 219-226.
- CONTOLI L., 1977 (a). Mammiferi del Tolfetano-Cerite (Lazio). In AA.VV., Problemi attuali di scienza e di cultura. Ricerche ecologiche , floristiche e faunistiche nel comprensorio Tolfetano- Cerite- Manziate. Accademia Nazionale dei Lincei, 227: 191- 226.
- CONTOLI L., 1977 (b). Problemi di gestione ambientale nel Tolfetano- Cerite- Manziate (Lazio). In AA.VV., Problemi attuali di scienza e di cultura. Ricerche ecologiche, floristiche e faunistiche nel comprensorio Tolfetano- Cerite- Manziate. Accademia Nazionale dei Lincei, 227: 267- 315.
- CORBETT, R.L., R.L. Marchinton & C.E. hill, 1971: Preliminary study of the effects of dogs on radio-equipped deer in a mountainous habitat. – Proc. S.E. Assoc. Game and Fish Comm. Conf. 25:69-77
- COX D.R., HINKLEY A.J.M., 1974. Theoretical statistics. Chapman and Hall, New York.
- CLUTTON-BROCK, T.H. & P.H. HARVEY. 1978. Mammals, resources and reproductive strategies. Nature, 273, 191-195
- DANILKIN A., DARMAN Y.A., MINAYEV A.N., SEMPERE A., 1995. Social structure, behaviour and ecological parameters of mygratory population of the Siberian roe deer (Capreolus pygargus). Transactions of the Academy of Sciences Biol. Ser., 1:48-61.
- Danilkin A. in association with HEWISON A.J.M., 1996. Behavioural ecology of siberian and european roe deer. Chapmann & Hall. London. 277pp.
- DE BOER H. Y., VAN BREUKELEN L., HOOTSMANS M.J.M. & WIEREN S.E., 2004. Flight distance in roe deer Capreolus capreolus and fallow deer Dama Dama as related to hunting and other factors. Wildlife Biology 10:1 (2004).
- DUNCAN P., TIXIER H., HOFMANN R.R., LECHNER-DOLL M., 1998. Feeding startegies and the physiology of digestion in roe deer. Andersen R., Duncan P., Linnel J. (Eds.): The European Roe Deer: the biology of success. Scandinavian Universoty Press. Oslo, 91-116.
- DEVOTO G., LOMBARDI G., 1977. Le formazioni sedimentarie ed eruttive del settore Tolfetano- Cerite- Manziate (Lazio Nordoccidentale). In AA.VV., Problemi attuali di scienza e di cultura. Ricerche ecologiche , floristiche e faunistiche nel comprensorio Tolfetano- Cerite- Manziate. Accademia Nazionale dei Lincei, 227: 5-35.
- DI CARLO A., 1977. L’avifauna del comprensorio Tolfetano-Cerite-Manziate (Lazio settentrionale). In AA.VV., Problemi attuali di scienza e di cultura. Ricerche ecologiche , floristiche e faunistiche nel comprensorio Tolfetano- Cerite- Manziate. Accademia Nazionale dei Lincei, 227: 125-176.
- DZIECIOLOWSKI R.,WASILEWSKI M. and PRZYPASNIAK J., 1990. Home ranger of roe deer (Capreolus capreolus) inhabiting a fine-grained landscape. Gibier Faune Sauvage, Game wild. Vol 15 (Hors serie Tome 2), p. 555-563.
- FARAGLIA E., RIGA F., 1997. Guida alla natura dei Monti della Tolfa. Muzzio Editore, Padova :1-223.
- FERIONI M., MERIGGI A., 1998. Selezione dell’habitat nel capriolo e modelli di valutazione ambientale in due aree dell’appennino pavese. II Congresso Italiano di Theriologia “I mammiferi in Italia:status, tendenze e implicazioni gestionali”. Varese 28-30 Ottobre: 117.
- FESTA E., 1925. Il Capriolo dell’Italia centrale.Boll. Museo Zool. An. Comp. Univ. Torino, 40:1-2.
- FOCARDI S., TOSO S., PAMPIRO F., RUIU P., PINTUS A., 1991. The diet of ungulates in a coastal mediterranean forest. In: Proceedings of the XXth Congress of the IUGB, 21-26 August 1991, 78-87.
- Gaillard J.M., Liberg O., Andersen R., Hewison A.J.M., Cederlund G., 1998. Population dynamics of roe deer. In: Andersen R., Duncan P., Linnel J. (Eds.): The European Roe Deer: the biology of success. Scandinavian University Press. Oslo, 309-336.
- Gerard J.F., Le Pendu Y., Maublanc M.L., Vincent J.P., Poulle M.L., Cibien C., 1995. Large group formation in european roe deer : an adaptive feature ? Revue d’ecologie (Terre Vie), 50: 391-401.
- Hewison A.J.M., Vincent J.P., Reby D., 1998. Social organisation of European roe deer. In: Andersen R., Duncan P., Linnel J. (Eds.): The European Roe Deer: the biology of success. Scandinavian University Press. Oslo, 189-221.
- HOOGE P. N. AND B. EICHENLAUB. 1997. Animal movement extension to arcview. ver. 1.1. Alaska Science Center – Biological Science Office, U.S. Geological Survey, Anchorage, AK, USA
- JEPPESEN J.L. (1990). Home range and movements of free-ranging roe deer (Capreolus capreolus) at Kalo. Dan. Rev. Game Biol.14 (1), pp.14.
- JEPPESEN J.L. 1987a. Impact of Human Disturbabance on home Range, Movements and Activity or Red Deer (Cervus elaphus) in a Danish Enviroment. Dan.Rev.Game Biol 13(2),38 pp.
- JEPPESEN J.L. 1984 : Human disturbance of foe deer and red deer: Preliminary results.- Commun. Inst. For. Fenn. 120: 113-118
- JEPPESEN J.L. 1987b . The disturbance effects of orienteering and hunting on roe deer (Capreolus capreolus). Dan.Rev.Game Biol 13(2),24 pp.
- JEWELL, .R.R. 1966. The concept of home range in mammals. Symp.Zool.Soc.Lond.No. 18, 85-109
- LINNEL J.D.C., DUNCAN P., ANDERSEN R., 1998. The European roe deer: a portrait of a successful species. In Andersen R., Duncan P., Linnel J. (Eds.): The European Roe Deer: the biology of success. Scandinavian Universoty Press. Oslo, 11-20.
- Linnel J.D.C., Wahlström K., Gaillard J.M., 1998. From birth to independence: birth, growth, neonatal mortality, hiding behaviour and dispersal. In: Andersen R., Duncan P., Linnel J. (Eds.): The European Roe Deer: the biology of success. Scandinavian University Press. Oslo, 257-285.
- Lister A.M., Grubb P., Sumner S.R.M., 1998. Taxonomy, morphology and evolution of European roe deer. In: Andersen R., Duncan P., Linnel J. (Eds.): The European Roe Deer: the biology of success. Scandinavian University Press. Oslo, 23-46.
- LORENZINI R.; BURRINI L.; MAZZONI DELLA STELLA R. (1997). Biochemical genetic differentiation in some roe deer populations of Tuscany, central Italy.Ital. J. Zool., 64: 239-244.
- LORENZINI R.; LOVARI S.; MASSETI M. (2002). The rediscovery of the Italian roe deer: genetic differentiation and management implications.Ital. J. Zool., 69: 367-379.
- KENWARD R., 1987. Wildlife Radio Tagging: equipment, field techniques and data analysis. Academic Press, London:1-222.
- KLEIN F. 1990. La reinroduction du cerf Cervus elaphus. Rev. Ecol, Suppl. 5.
- KOUBEK P., 1995. Home range dynamics and movements of roe deer (Capreolus capreolus) in a floodplain forest. Folia Zoologica- 44(3) 215-226.
- KRAMER D.L. e BONEFANT M., 1997. Direction of predator approach and the decision to flee to a refuge. Anim. Behav.,54,289-295
- MACARTHUR R. A., V. GEIST, R. H. Johnston, 1982 – Cardia and behavioural rsponses of mountain sheep: a physiological approach to the study of wildlife harassment. Can. J. Zool., 57: 2010-2021
- MARCHAL C., GERARD J.F., DELORME D., BOISAUBERT B. and BIDEU E.,1998. Space and habitat use by field roe deer (Capreolus capreolus) in mid winter and mid-growing season. Giber Faune Sauvage, Game wildl. Vol 15, p 737-746.
- MARCONE F., 2003. Tesi di Laurea, Università degli studi di Roma “La Sapienza”.
- MCLAREN M., J. E. GREEN, 1985 – The reactions of muskoxen to snowmobile harassment. Arctic, 38: 188-193
- MILLSPAUGH J.J., MARZLUFF J.M., 2001. Radio Tracking and Animal Populations. Academic Press, London: 1-473.
- MONTELUCCI G.,1977. Note preliminari sulla flora e sulla vegetazione delle cerrete di Manziana e di Canale-Monterano. In AA.VV., Problemi attuali di scienza e di cultura. Ricerche ecologiche , floristiche e faunistiche nel comprensorio Tolfetano- Cerite- Manziate. Accademia Nazionale dei Lincei, 227: 51-74.
- MYSTERUD A., 1999.Seasonal migration pattern and home range of roe deer (Capreolus capreolus) in an altitudinal gradient in southern Norway. J. Zool. Lond., 247: 479-486.
- MYSTERUD A., IMS R.A., 1998. Functional responses in habitat use: availability influences relative use in trade-off situations. Ecology, 79(4): 1435-1441.
- NAMS V.O., 1990. Locate II user’s guide. Pacer Computer software, Truro. 70pp.
- PATTERSEN I.J., 1988 – Response of Appendine Chamois to human disturbance. Z. Saugetierk., 53: 245-252
- PEDROTTI L., TOSI G., FACOETTI R., PICCININI S., 1995. Organizzazione di uno studio mediante radio-tracking e analisi degli home-range: applicazione agli Ungulati alpini. In: Applicazioni del radio-tracking per lo studio e la conservazione dei vertebrati. Suppl. Ric. Biol. Selvaggina Vol. 23: 3-100.
- PERCO F., 1987. Ungulati. Editore Lorenzini, Udine.
- PERCO F., CALO’ C.M., 1994. The status of roe deer in Italy. In: Wotschikowsky U. (Ed.): Proceedings of the 2nd European Roe Deer meeting.Wildbiologische Gesellschaft. Munchen, pp 43-53.
- PERCO F., PERCO D.,1979. Il capriolo. Ed. Carso, Trieste, 211pp.
- PIAZZI A., 2002. Tesi di Laurea, Università degli studi di Roma “La Sapienza”.
- PICCININI S., OREILLER P., DOMENEGHETTI L., VIGLINO P., 1998. Risultati di immissione di individui di capriolo provenienti da stabulazione in centro di recupero e monitorati con tecniche di radio-tracking. II Congresso Italiano di Theriologia “I mammiferi in Italia:status, tendenze e implicazioni gestionali”. Varese 28-30 Ottobre: 114.
- Randi E., Pierpaoli M., Danilkin A., 1998. Mitochondrial DNA polymorphism in population of Siberian and European roe deer (Capreolus pygargus and C. capreolus). In: Heredity, 80: 429-437.
- RANDI E., MUCCI N., 2001. Genetic structure and origin of roe deer populations in Italy. 5th European Roe Deer Meeting, Tredozio (FO) 24-27 Aprile.
- RETTIE W.J., McLOUGHLIN P.D., 1999. Overcoming radiotelemetry bias in habitat-selection studies. Canadian Journal of Zoology, 77: 1175-1183.
- RICHENS V. B., G. R. LAVIGNE, 1978 – Response of white-tailed deer to snowmobiles and snowmobiles trails in Maine. Can. Field-Nat.,92: 334-344.
- ROOT B.G. , FRITZELL E.K. and GIESSMAN N.F, 1988. Effects of intensive hunting on white-tailed deer movement. Wild. Soc. Bull. 16:145-151.
- SALTZ D., 1994. Reporting error measures in radio location by triangulation: a review. Journal of Wildlife Management, 58(1): 181-184.
- San Josè C., Lovari S., 1998. Ranging movements of female roe deer: do home-loving does roam to mate? Ethology, 104: 721-728.
- San Josè C., Lovari S., Ferrari N., 1997. Grouping in roe deer: an effect of habitat openness or cover distribution? Acta Theriologica, 42: 235-239.
- SANDERSON, G.C. 1966 The study of mammal movements – a review. J. Wildl.Mgmt. 30(1), 215-235
- SCHONFELDER I., SCHONFELDER P., 1998. La flora mediterranea. DeAgostini, Novara: 1-320.
- SCHWAB G., 1994. Home-ranges and habitat use of the Hahnebaum roe deer. In: Wotschikowsky U. (Ed.): Proceedings of the 2nd European Roe Deer meeting.Wildbiologische Gesellschaft. Munchen, 123-128.
- SERRAO G., 2004. Tesi di Laurea, Università degli studi di Roma “La Sapienza”.
- SHULTZ R. D., J. A. BAILEY, 1978 – Response of national park elk to human activity. J. Wildl. Manage., 42:91-100
- SONETTI M.G. E GALIMBERTI F., 1991. Pubblicazione interna all’I.N.F.S Documenti Tecnici.
- SPADA F, 1977. Primi lineamenti della vegetazione del comprensorio Tolfetano-Cerite. In AA.VV., Problemi attuali di scienza e di cultura. Ricerche ecologiche , floristiche e faunistiche nel comprensorio Tolfetano- Cerite- Manziate. Accademia Nazionale dei Lincei, 227: 37-50.
- STRANDGAARD H., 1972. The roe deer (Capreolus capreolus) population at Kalo and the factors regulating its size. Danish Review of game biology. Vol. 7(1) pp 205.
- SWESON J.E., 1982. effects of hunting on habitat use by mule deer on mixed-grass prairie in montana. Wildl. Soc. Bull. 10: 115-120
- SWIHART R.K., SLADE N.A., 1985. Influence of sampling interval on estimates of home-range size. J. Wildl. Manage., 49(4): 1019-1025.
- TIXIER H., DUNCAN P., 1996. Are European roe deer browsers? A review of variation in the composition of their diet. Revue d’Ecologie (Terre Vie) 51:3-17.
- TASSI F. (1976)., 1 La reintroduzione degli ungulati nell’Appennino centrale. Contributi scientifici alla conoscenza del parco d’Abruzzo .
- TOSI G., TOSO S., 1992. Indicazioni generali per la gestione degli ungulati. I.N.F.S., Documenti Tecnici n.11: 3-144.
- TOSO S., 2001. Roe deer in Italy: distribution, status and management. 5th Roe deer Meeting, Tredozio (FO) 24-27 Aprile.
- TUFTO J.; ANDEREI R.; LINNELL J. (1996). Habitat use and ecological correlates of home range size in a small cervid: the roe deer. Journal of Animal Ecology, 65: 715-724.
- Vincent J.P., Bideau E., Hewison A.J.M., Angibault J.M., 1995. The influence of increasing density on body weight, kid production, home range and winter grouping in roe deer (Capreolus capreolus). Journal of Zoology, 236: 371-382.
- VIRGOS E., TELLERIA J.L., 1998. Roe deer habitat selection in Spain: constrains on the distribution of a species. Canadian Journal of Zooog. 76:1294-1299.
- WHITE G.C. e GARROTT R.A., 1990. Analysis of wildlife radiotracking data. Academic Press, San Diego. pp 383.
Pubblicato il
19 febbraio 2014




